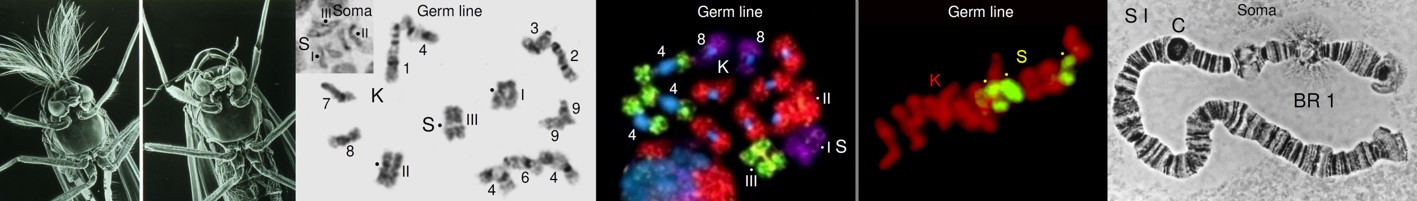
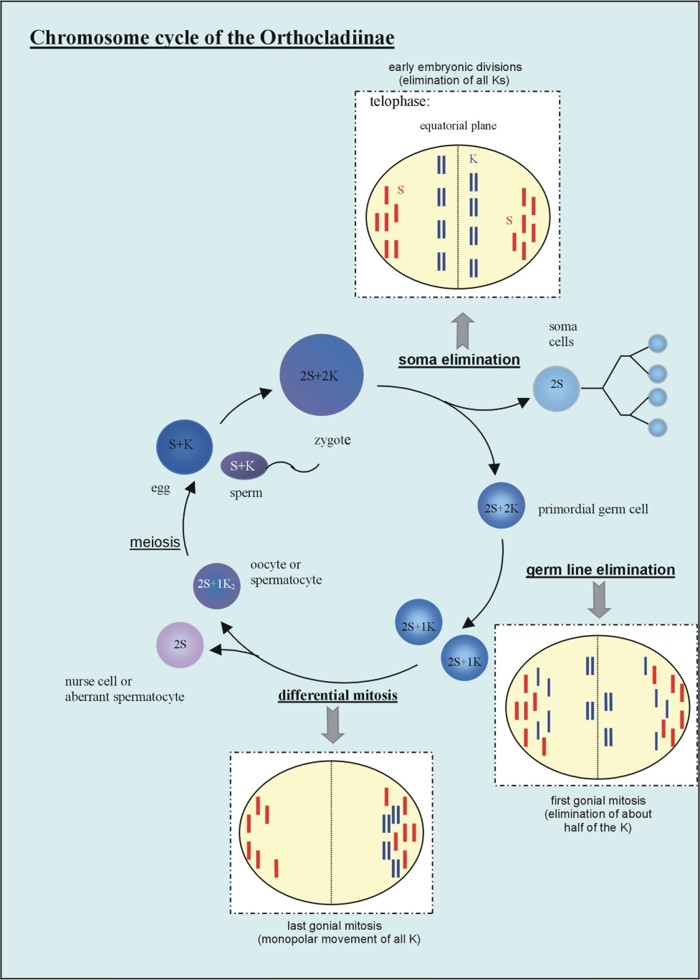
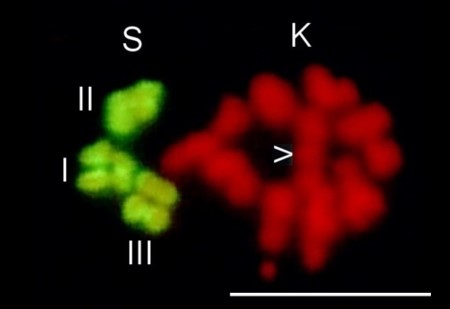
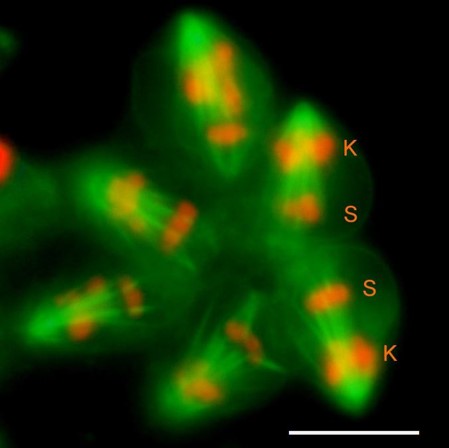
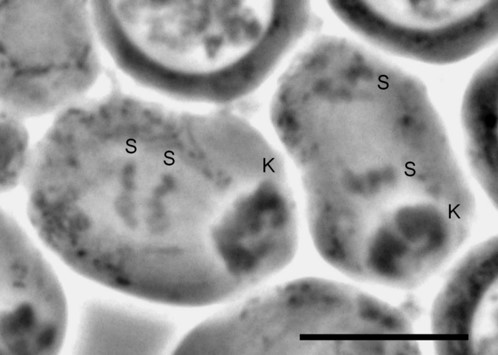
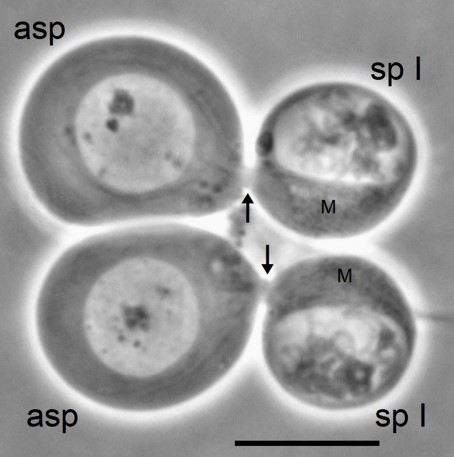
Fig. 6 Diagram summarising the most relevant chromosomal events in male differential gonial mitosis, meiosis I and meiosis II of Acricotopus lucidus. To clarify interpretation, both chromatids are represented in the condensed chromosomes. S, soma chromosomes (red; 2n=6); K, germline-limited chromosomes (blue; variable number); asp, aberrant spermatocyte; sp I, primary spermatocyte; sp II, secondary spermatocyte; sp, spermatid.
These events indicate a programmed inactivation of all chromosomes in the aberrant spermatocyte at the beginning of meiosis. The inactivation of the Ss is suggested to have developed during evolution to inhibit the entry of the aberrant spermatocytes into meiosis, thereby preventing the formation of sperms containing only Ss but no Ks (Staiber 2016).
References
Bauer, H (1970) Rearrangements between germ-line limited and somatic chromosomes in Smittia parthenogenetica (Chironomidae, Diptera). Chromosoma 32:1-10
Bauer, H, Beermann, W (1952) Der Chromosomencyclus der Orthocladiinen (Nematocera, Diptera). Z Naturforsch 7b:557-563
Beermann, W (1956) Nuclear differentiation and functional morphology of chromosomes. Cold Spring Harbor Symp Quant Biol 21:217-230
Staiber W (2007). Asymmetric distribution of mitochondria and of spindle microtubules in opposite directions in differential mitosis of germ line cells in Acricotopus. Cell Tissue Res 329:197–203
Staiber W (2008) Centrosome hyperamplification with the formation of multiple asters and programmed chromosome inactivation in aberrant spermatocytes during male meiosis in Acricotopus. Cell Tissue Res 334:81-91
Staiber, W (2016) Loss of centromeric histone H2AT120 phosphorylation accompanies somatic chromosomes inactivation in the aberrant spermatocytes of Acricotopus lucidus (Diptera, Chironomidae). Protoplasma 253:211-216
White, MJD (1973) Animal cytology and evolution, 3rd edn, Cambridge University Press, Cambridge. pp 500-546